Augmentons la diversité florale des milieux pour une meilleure cohabitation entre pollinisateurs sauvages et abeilles domestiques !
A l'occasion de la Journée mondiale de l'abeille qui se tient ce 20 mai, Noé vous propose de revenir sur un sujet fondamental : la cohabitation entre les insectes pollinisateurs sauvages et les abeilles domestiques, grâce à une plus grande diversité de fleurs pour l'ensemble de ces espèces.
Rappelons que le vivant dépend des pollinisateurs et qu'il est donc crucial d'enrayer leur déclin et de freiner la perte de biodiversité et la dégradation des écosystèmes. La pollinisation est un processus fondamental pour la survie des écosystèmes car de lui dépendent la reproduction de près de 90% des plantes sauvages à fleurs, 75% des cultures vivrières et 35% des terres agricoles à l’échelle de la planète.
Les pollinisateurs sauvages se comptent par milliers…
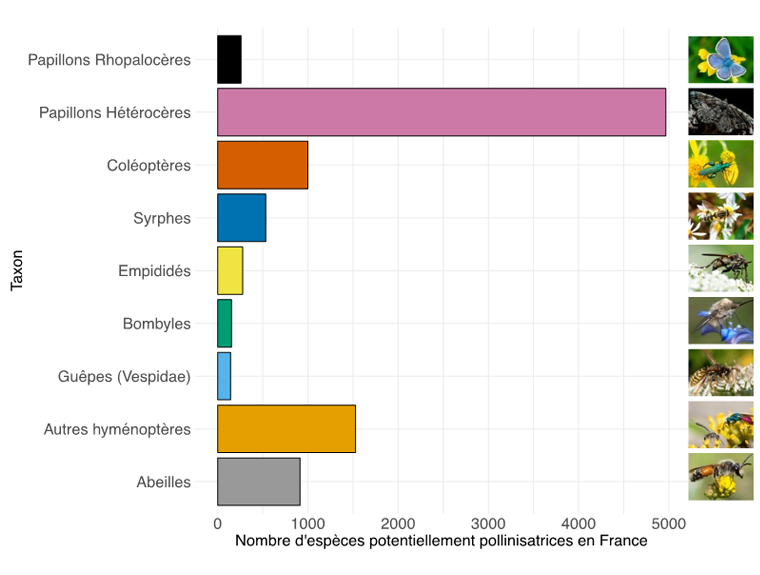
Les pollinisateurs sauvages rassemblent plusieurs milliers d’espèces en France, dont environ 1000 espèces d’abeilles ainsi que de nombreux autres groupes taxonomiques (résumés en figure 1).
Figure 1 : Nombre d’espèces potentiellement présentes en France1. Les papillons Rhopalocères correspondent aux papillons de jours, les papillons Hétérocères aux papillons de nuit. Les syrphes, empididés et bombyles sont des « mouches ». Les hyménoptères forment l’ordre auxquelles appartiennent les guêpes et abeilles.
DES INSECTES QUI varient énormément dans leur écologie…
Ils ont notamment des habitats très variables selon les espèces. Certaines espèces sont par exemple inféodées aux végétations herbacées des milieux ouverts comme les prairies permanentes, ou d’autres aux forêts, ce qui peut avoir des conséquences déclinleurs effectifs et donc leur statut de conservation.
Par exemple, la perte des prairies permanentes a un impact particulièrement important sur des espèces d’abeilles solitaires et spécialisées comme Andrena hattorfiana et Andrena humilis en Suède (où 97 à 99% des prairies gérées ont disparu au cours des dernières décennies)2.
...et quant à leur phénologie
Ils ne sont pas actifs aux mêmes périodes de l’année, ni même de la journée (une partie des pollinisateurs n’étant même actifs que la nuit). Ainsi, il existe une forme de répartition de leur activité dans le temps qui induit un certain niveau de complémentarité (voir ci-dessous) entre pollinisateurs 3, 4.
la complémentarité fonctionnelle, qu’est-ce que c’est ?
Prenons l’exemple simplifié de la profondeur des corolles et de la longueur de la langue des insectes.
Imaginons une prairie dans laquelle on ne trouve que des fleurs avec des corolles peu profondes, avec un cortège de pollinisateurs composé d’un groupe avec une langue longue, et d'un autre groupe avec une langue courte. Les pollinisateurs avec une langue longue vont être moins efficaces pour collecter du nectar que les espèces avec des pièces buccales courtes5. La compétition va donc être forte, et en défaveur des espèces les moins adaptées à la profondeur des corolles.
Si par contre, dans cette prairie on trouve à la fois des fleurs avec des corolles profondes et des corolles peu profondes, les pollinisateurs à langue longue seront plus efficaces sur les fleurs à corolle profonde et auront donc plus tendance à les exploiter pour optimiser leur niveau de récompense, alors que les pollinisateurs à langue courte auront tendance à rester sur les corolles peu profondes pour les mêmes raisons.
Il y a donc complémentarité entre les deux groupes de pollinisateurs : plus des espèces occupent des niches complémentaires, moins elles sont en compétition les unes avec les autres6.
Cependant, la niche ne se résume pas à la seule longueur de langue en interaction avec la profondeur de la corolle : de nombreuses autres caractéristiques ou traits entrent en jeu ! Et pour prendre en considération ces autres traits, on utilise des indices de diversité fonctionnelle qui permettent, entre autres, d’évaluer à quel point au sein d’un type de milieu donné, les espèces de plantes et de pollinisateurs présents ont des caractéristiques qui se chevauchent ou se complètent. Si le chevauchement est fort, il y a redondance fonctionnelle entre les espèces. Si le chevauchement est faible, il y a complémentarité7 !
Des insectes qui n’ont pas non plus les mêmes besoins alimentaires
Certains, comme les abeilles, collectent pollen et nectar alors que les lépidoptères (papillons) adultes consomment essentiellement du nectar car leurs larves se nourrissent par elles-mêmes sur les tissus des plantes ou des lichens.
Ils n’ont par ailleurs pas accès aux mêmes ressources alimentaires que ce soit lié à la longueur de leurs pièces buccales mais également à l’anatomie de ces pièces qui conditionnent leur efficacité à prélever du nectar selon sa concentration en sucre et donc sa viscosité.
Par exemple, les lépidoptères avec leur trompe, prélèvent le nectar par capillarité et sont donc plus efficaces à prélever ce dernier sur des fleurs possédant un nectar peu visqueux. Les diptères possédant des pièces buccales de type lécheur (par exemple les syrphes), ont une préférence sur les fleurs produisant un nectar visqueux et facile d’accès (typiquement des plantes de la famille des apiacées). Ces différents critères conditionnent donc les plantes avec lesquelles les pollinisateurs interagissent, mais également avec quels autres pollinisateurs ils vont entrer en compétition ainsi que leur propension à être impactés par des pressions anthropiques qui auraient par exemple pour effet de modifier la composition de la végétation8 !
Le déclin des pollinisateurs sauvages est lié aux pressions anthropiques
Et ce sont justement ces pressions anthropiques qui sont principalement à l’origine du déclin des pollinisateurs sauvages, cela inclut la destruction de leurs habitats par l’intensification de l’agriculture et l’artificialisation des sols, les différentes formes de pollution (ex : pollution par les produits phytopharmaceutiques, pollution lumineuse), et d’autres facteurs comme le changement climatique9.
Ce déclin des pollinisateurs est d'ailleurs très médiatisé (la journée de l’abeille en étant un exemple parmi tant d’autres), mais avec un biais encore bien présent.
En effet, l’abeille domestique est bien plus l’objet de la sympathie du grand public que les autres pollinisateurs. C'est donc préférentiellement en faveur de l’abeille domestique que de nombreuses actions sont ainsi entreprises (parrainage de ruches par les particuliers ou les entreprises, implantation massive de ruchers en ville) sous couvert d’actions en faveur des pollinisateurs ou de la biodiversité10.
Bien que l’apiculture soit un moyen potentiellement efficace pour faire découvrir aux citoyens les pollinisateurs et les périls auxquels ils font face, cette mode de l’apiculture urbaine demeure défavorable aux pollinisateurs sauvages présents dans ces villes, avec une compétition avérée pour les ressources florales entre une partie de ces insectes et l’abeille domestique11. Plus généralement, de plus en plus d’études soulignent une compétition entre abeille domestique et pollinisateurs sauvages, que ce soit en milieu urbain ou en milieu agricole même lorsque des bandes fleuries sont implantées localement, mais également dans des aires protégées12–15.
Comment expliquer ce phénomène de compétition ?
Pour expliquer ce phénomène de compétition, revenons à nos traits et au phénomène de complémentarité présentés précédemment. En effet, dans une récente étude issue des travaux d’une équipe de chercheurs italiens16, ces derniers ont observé que plus les pollinisateurs sauvages, et en particulier les abeilles sauvages, ont une longueur de langue équivalente à celle de l’abeille domestique, plus ils sont en compétition avec elle à cause d’un fort niveau de recouvrement de niche.
Cependant, ces mêmes chercheurs ont identifié qu’une augmentation de la diversité fonctionnelle des traits floraux (taille des fleurs et couleur dans cette étude) d’un habitat (ici des prairies) diminuait la compétition entre abeille domestique et pollinisateurs sauvages en permettant aux pollinisateurs sauvages d’exploiter d’autres ressources disponibles.
Implanter des habitats diversifiés pour enrayer les déclin des pollinisateurs
Ainsi, pour permettre une meilleure coexistence entre pollinisateurs sauvages et apiculture, il est indispensable d’agir pour implanter et restaurer des habitats à la fois fortement diversifiés en termes d'espèces végétales, mais également en termes de traits floraux caractéristiques, et ce à la fois à des échelles locales (ex : une parcelle de prairie, une bande enherbée) jusqu’à des échelles paysagères.
Parmi ces habitats, les prairies permanentes tiennent une place centrale alors qu’environ 3 millions d’ha d’entre elles ont été perdus entre 1960 et 2010 en France (soit une perte d’environ 30% de la surface en prairies permanentes) et que les prairies restantes sont majoritairement dans un état de conservation défavorable à la biodiversité. Pour favoriser la diversité des ressources florales, il est indispensable d’aller bien au-delà des seules jachères fleuries annuelles ou des bandes de luzerne ! Il est également indispensable d’agir avec les acteurs des autres filières animales concernées (bovins, ovins…) et de lutter contre l’artificialisation des sols.
Cette diversification des habitats est essentielle pour la conservation des pollinisateurs sauvages, mais également pour l’apiculture. Par exemple, la diversité de la ressource en pollen est un facteur clef de santé d’une colonie d’abeilles domestiques ou du succès rencontré dans le cadre de l’élevage de reine. Les apiculteurs, dans un cadre sanitaire de plus en plus incertain avec l’arrivée successive concrète ou possible d’espèces impactant leur production (Varroa destructor, Aethina tumida, Tropilaelaps…) ont donc un intérêt réel à s’engager dans la restauration ou l’implantation massive de ces habitats !
Ainsi, plutôt que de fuir les milieux agricoles pour se réfugier dans les villes ou les aires protégées menant à une régulation légitime qui pourrait prendre de multiples formes17, plutôt que de privilégier un solutionnisme court-termiste à coût d’essences d’arbres et de lianes exotiques18, plutôt que de devoir nourrir ses ruches au sucre industriel (dont sucre de betterave, seule culture en France bénéficiant d’une dérogation pour l’utilisation de néonicotinoïdes malgré la persistance de ce produit dans le sol et donc dans les cultures de la rotation, dont le colza et ses fleurs19) pour passer les disettes qui frappent les paysages appauvris en ressources florales, les apiculteurs ont un intérêt réel à lutter conjointement avec les associations de protection de l’environnement contre l’agriculture intensive et l’urbanisation, les deux principaux vecteurs de destruction des habitats des pollinisateurs sauvages et de perte de ressources florales pour une large partie des pollinisateurs. Apiculteurs et « environnementalistes » ont donc des intérêts communs et doivent agir conjointement afin d’enrayer le déclin de ces formidables insectes !
Sources et références :
1 S. Gadoum and J.-M. Roux-Fouillet, ‘Plan national d’actions « France Terre de pollini- sateurs » pour la préservation des abeilles et des insectes pollinisateurs sauvages’, Office Pour les Insectes et leur Environnement – Ministère de l’Écologie, du Développement Durable et de l’Énergie, 2016.
2 A. Nieto et al., European red list of bees. Luxembourg: Publications Office, 2014. Accessed: May 20, 2020. [Online]. Available: http://bookshop.europa.eu/uri?target=EUB:NOTICE:KH0714078:EN:HTML
3 C. Venjakob, A.-M. Klein, A. Ebeling, T. Tscharntke, and C. Scherber, ‘Plant diversity increases spatio-temporal niche complementarity in plant-pollinator interactions’, Ecol. Evol., vol. 6, no. 8, pp. 2249–2261, Apr. 2016, doi: 10.1002/ece3.2026.
4 R. E. Walton, C. D. Sayer, H. Bennion, and J. C. Axmacher, ‘Nocturnal pollinators strongly contribute to pollen transport of wild flowers in an agricultural landscape’, Biol. Lett., vol. 16, no. 5, p. 20190877, May 2020, doi: 10.1098/rsbl.2019.0877.
5 S. G. T. Klumpers, M. Stang, and P. G. L. Klinkhamer, ‘Foraging efficiency and size matching in a plant–pollinator community: the importance of sugar content and tongue length’, Ecol. Lett., vol. 22, no. 3, pp. 469–479, Mar. 2019, doi: 10.1111/ele.13204.
6 J. Fründ, C. F. Dormann, A. Holzschuh, and T. Tscharntke, ‘Bee diversity effects on pollination depend on functional complementarity and niche shifts’, Ecology, vol. 94, no. 9, pp. 2042–2054, Sep. 2013, doi: 10.1890/12-1620.1.
7 N. W. H. Mason, D. Mouillot, W. G. Lee, and J. B. Wilson, ‘Functional richness, functional evenness and functional divergence: the primary components of functional diversity’, Oikos, vol. 111, no. 1, pp. 112–118, Oct. 2005, doi: 10.1111/j.0030-1299.2005.13886.x.
8 J. Goulnik, S. Plantureux, I. Dajoz, and A. Michelot-Antalik, ‘Using Matching Traits to Study the Impacts of Land-Use Intensification on Plant–Pollinator Interactions in European Grasslands: A Review’, Insects, vol. 12, no. 8, p. 680, Jul. 2021, doi: 10.3390/insects12080680.
9 P. Eggleton, ‘The State of the World’s Insects’, Annu. Rev. Environ. Resour., vol. 45, no. 1, p. annurev-environ-012420-050035, Oct. 2020, doi: 10.1146/annurev-environ-012420-050035.
10 B. Geslin, L. Ropars, M. Zakardjian, and F. Flacher, ‘The misplaced management of bees’, Preprints, preprint, Jan. 2022. doi: 10.22541/au.164319695.57033003/v1.
11 L. Ropars, I. Dajoz, C. Fontaine, A. Muratet, and B. Geslin, ‘Wild pollinator activity negatively related to honey bee colony densities in urban context’, PLOS ONE, vol. 14, no. 9, p. e0222316, Sep. 2019, doi: 10.1371/journal.pone.0222316.
12 L. Ropars et al., ‘Land cover composition, local plant community composition and honeybee colony density affect wild bee species assemblages in a Mediterranean biodiversity hot-spot’, Acta Oecologica, vol. 104, p. 103546, Apr. 2020, doi: 10.1016/j.actao.2020.103546.
13 L. Ropars, L. Affre, E. Thébault, and B. Geslin, ‘Seasonal dynamics of competition between honey bees and wild bees in a protected Mediterranean scrubland’, Oikos, vol. 2022, no. 4, 2022, doi: https://doi.org/10.1111/oik.08915.
14 V. A. Wojcik, L. A. Morandin, L. Davies Adams, and K. E. Rourke, ‘Floral Resource Competition Between Honey Bees and Wild Bees: Is There Clear Evidence and Can We Guide Management and Conservation?’, Environ. Entomol., vol. 47, no. 4, pp. 822–833, Aug. 2018, doi: 10.1093/ee/nvy077.
15 G. M. Angelella, C. T. McCullough, and M. E. O’Rourke, ‘Honey bee hives decrease wild bee abundance, species richness, and fruit count on farms regardless of wildflower strips’, Sci. Rep., vol. 11, no. 1, p. 3202, Dec. 2021, doi: 10.1038/s41598-021-81967-1.
16 A. Cappellari, ‘Functional traits of plants and pollinators explain resource overlap between honeybees and wild pollinators’, p. 11, 2022.
17 M. Henry and G. Rodet, ‘The apiary influence range: A new paradigm for managing the cohabitation of honey bees and wild bee communities’, Acta Oecologica, vol. 105, p. 103555, May 2020, doi: 10.1016/j.actao.2020.103555.
18 Y. Darricau, Planter des arbres pour les abeilles - L’api-foresterie de demain, Terran Eds De. 2018.
19 D. Wintermantel, J.-F. Odoux, A. Decourtye, M. Henry, F. Allier, and V. Bretagnolle, ‘Neonicotinoid-induced mortality risk for bees foraging on oilseed rape nectar persists despite EU moratorium’, Sci. Total Environ., vol. 704, p. 135400, Feb. 2020, doi: 10.1016/j.scitotenv.2019.135400.